Suplement CLXXXIII
ZESZYTY NAUKOWE UNIWERSYTETU JAGIELLOŃSKIEGO NR 23 PRACE ZOOLOGICZNE 1959. Z. 4, SS. 133-149.
Stefan Gut
Nitkowate promienie płetwowe u ryb z rodzaju Colisa i Trichogaster
jako narządy zmysłowe dotykowo-chemiczne
The pectoral filaments in the fishes of genera Colisa and Trichogaster
as organs of the feeling and chemical sense
Zakład Psychologii i Etologii Zwierząt UJ w Krakowie
Kierownik: prof dr R. J. Wojtusiak
Wstęp
Celem niniejszej pracy było zbadanie funkcji zmysłowej nitkowatych promieni płetw piersiowych występujących u ryb z rodzaju Colisa i Trichogaster z rodziny Osphromenidae, z rzędu Labyrinthici. Ponieważ ryby te wyrostkami swymi obmacują przestrzeń przed i pod sobą przy omijaniu rozmaitych przeszkód oraz różne ciała, wśród nich także pokarm przed uchwyceniem go pyszczkiem, nasuwało się przypuszczenie, że nitkowate promienie płetwowe mogą służyć u nich jako narządy dotykowe lub chemiczne. Podstawę do przypuszczenia tego dawał fakt, że u ryb żerujących na mulistym dnie zbiorników wodnych istnieje zwyczaj wyszukiwania pokarmu przy pomocy różnego rodzaju wąsów lub promieni skrzelowych.
Zwierzęta chwytają wszelkie substancje pokarmowe, które znajdą się w kontakcie z tymi wyrostkami, pomijając inne nie nadające się na pokarm. Herrick (1903) zauważył, że Microgadus poszukujący pokarmu, ciągnie swe nitkowate wyrostki po dnie. Ponieważ natrafia tymi utworami płetw brzusznych na odpowiednie ciała, gdy głowa je już minęła, przeto aby połknąć pokarm, musi się nieco cofnąć. U ryb poszukujących pokarmu przy pomocy wyrostków wysuwanych ku przodowi, jak u przedstawicieli rodzajów Colisa i Trichogaster, pyszczek znajduje się w chwili zetknięcia się wyrostków z pokarmem przed lub nad nim i ryby reagują na pokarm skokiem w przód lub w dół. U ryb z rodzaju Trigla znane były w górnej części rdzenia przedłużonego trzy pary zgrubień, lobi accessorii. Ussow (1882) stwierdził, że wybiegające z nich nerwy unerwiają promienie płetw piersiowych. Bloch (1783) uważał je za wyrostki służące do przywabienia zdobyczy, natomiast Tiedemann (1813) za narządy ruchowe i dotykowe. Allen (1855) u gatunku Prionotus palmipes, spokrewnionym z Trigla, stwierdził, że wolne promienie płetwowe służą do grzebania w piasku, ruchu naprzód i jako narządy dotykowe. Jourdan (1890) u Peristedion cataphractum z tej samej rodziny Triglidae, nie znalazł w nabłonku promieni płetwowych guzków smakowych i wobec tego przypisał im jedynie rolę narządów dotykowych. Również Morril (1895) i Herrick (1904) odmówili wyrostkom tym roli narządów chemicznych. Dopiero Scharrer (1935) wyjaśnił ich funkcje u Trigla jako narządów dotykowych i chemicznych, które przy pełzaniu tej ryby po dnie i obmacywaniu napotkanych przedmiotów, informują zwierze o przydatności ich jako pokarmu.
Wrażliwość ryb na bodźce dotykowe i chemiczne była przedmiotem licznych badań. Zauważono wzmożoną aktywność ryb, gdy pokarm znajduje się w wodzie w pewnej od nich odległości. Niektórzy badacze, jak Nagel (1894) i Pieron (1908) główną rolę w odszukiwaniu pokarmu przypisywali jednak wzrokowi. Parker (1911, 1922) wykazał, że sumy amerykańskie z dwóch podanych im identycznych woreczków atakują jedynie ten, w którym jest pokarm. Na sole, kwasy i zasady wrażliwa jest u nich cała powierzchnia ciała. Herrick (1904) zauważył, że ryby zjadają kawałki mięsa, natomiast nie tykają innych przedmiotów podobnych do pokarmu z kształtu, wielkości i koloru. Zależnie od sposobu działania bodźca chemicznego, z bezpośredniej bliskości lub z odległości oraz od unerwienia i budowy danego narządu zmysłowego, wyróżniono u ryb osobne zmysły smaku i węchu (v. Frisch 1920, Strieck 1924, Krinner 1935, Warden, Jenkins, Warner 1936). Narządy smakowe zawierające zwykłe kubki smakowe, rozmieszczone są głównie w okolicy gębowej, głowy, na płetwach i innych miejscach ciała i unerwione są przez VII nerw (Olmsted 1920). Narządy węchowe zaopatrzone głównie w dołki węchowe, unerwione przez nerwy węchowe, występują w okolicy pyska, skrzel lub w pewnej odległości od pyska i czasem w pobliżu narządów smakowych, przez co trudno je od tych ostatnich odróżnić. Po przecięciu nerwów węchowych odnośne narządy przestają reagować na pokarm z odległości (Steiner 1885, Uexküll 1895, Sheldon 1909, 1911, Parker 1922), natomiast po przecięciu nerwu VII zanikają reakcje smakowe (Olmsted 1920). Strieck (1924) metodą tresury przekonał się, że strzeble (Phoxinus laevis) odróżniają smakowo pokarm od nitek bawełnianych podobnych do pokarmu. Według Parkera (1922) także wolne zakończenia nerwowe rozsiane po całej skórze działają jako receptory chemiczne, niezależnie, od tego czy unerwiają je nerwy węchowe czy smakowe.
Po wyeliminowaniu wpływu zakończeń nerwowych dotykowych i linii nabocznej przez przecięcie nerwu twarzowego (n. facialis) obsługującego je, ryby reagowały na mięso. Długotrwałe działanie prądu wody może zmęczyć zakończenia nerwów dotykowych tak, że zwierzę przestaje reagować na bodźce mechaniczne, natomiast normalnie reaguje na bodźce chemiczne. Ten sam wynik daje zastosowanie 2% roztworu kokainy, który eliminuje na przeciąg 10 — 20 minut reakcje dotykowe, natomiast nie usuwa wrażliwości na bodźce chemiczne. W niniejszej pracy rozpoznanie zmysłu dotykowego i chemicznego umożliwiło zastosowanie odpowiednich doświadczeń, opisanych poniżej.
Ryc. 1. Trichogaster leeri (Bleeker).
Wobec znacznej trudności przeprowadzenia ścisłego rozdziału między zmysłem węchu i smaku u ryb zamieszkujących płynny ośrodek, nie analizowano jakościowych różnic we wrażliwości odpowiednich zmysłów, lecz podporządkowano wszystkie reakcje ogólnemu pojęciu zmysłu chemicznego. Na podstawie danych z literatury, a zwłaszcza ze względu na unerwienie nitkowatych wyrostków skrzelowych przez nerw VII, można by je uważać za narząd smakowy, który jednak działa nie tylko w bezpośrednim kontakcie z danym ciałem, ale także umożliwia prawdopodobnie percepowanie odpowiednich substancji rozpuszczonych w wodzie.
Do badań użyto następujących gatunków i odmian ryb z rodziny Osphromenidae, żyjących w wodach płynących i stojących południowo-wschodniej Azji:
Colisa (Trichogaster) fasciata Bloch et Schneider
Colisa (Trichogaster) labiosa Day
Trichogaster leeri Blecker (ryc. 1)
Trichogaster (Trichopodus) trichopterus Pall. var. koelretueri Cuv. et Val.
(Osphromenus trichopterus) (ryc. 2)
Trichogaster trichopterus Pall. var. sumatranus Ladiges
Ryc. 2. Trichogaster trichopterus (Pallas.)
Mogą one żyć w wodach, które dla innych ryb byłyby zabijające z powodu małej ilości tlenu dzięki temu, że posiadają labirynt i mogą pobierać tlen z powietrza (Labyrinthici).
Doświadczenia przeprowadzano z wszystkimi wymienionymi formami, w tekście jednak i na tabelach podano dla przykładu dane odnoszące się do jednego lub dwu gatunków, gdyż nie zaobserwowano wyraźniejszych różnic w zachowaniu się poszczególnych gatunków czy odmian.
Doświadczenia wstępne
Dla przekonania się, czy nitkowate wyrostki płetwowe u ryb z rodziny Osphromenidae odgrywają jakąś rolę przy wyszukiwaniu pokarmu, przeprowadzono najpierw serię obserwacji wstępnych. Polegały one na tym, że na cienkim drucie podawano rybom żywego wazonkowca (Enchytreus) w ten sposób, aby ryba nie mogła dostrzec pokarmu przy pomocy wzroku, lecz aby go mogła dotknąć nitkowatym promieniem płetwy piersiowej. Z chwilą dotknięcia nim wazonkowca zwierzę zwracało się powoli w kierunku pokarmu. Gdy próby te powtarzano, reakcja ta następowała coraz wyraźniej. Rodzaj reakcji zależał także od ułożenia nitkowatych wyrostków. Jeżeli ułożone były one w chwili dotknięcia ich wazonkowcem wzdłuż ciała, następowało cofnięcie się ryby. Jeżeli wyrostki nitkowate wysunięte były naprzód, następował szybki ruch do przodu, chwyt za pokarm i połknięcie go. Wyniki doświadczeń tych, przeprowadzonych na gatunku Trichogaster trichopterus var. sumatranus, okazach o normalnie rozwiniętych oczach, przedstawione zostały na tabeli 1. Widać z nich, że ryby w 100% reagowały pozytywnie na dotknięcie wyrostków nitkowatych żywym wazonkowcem (Enchytreus).

Podobne reakcje można było zauważyć u ryb wyszukujących samodzielnie pokarm. Jeżeli został on wrzucony na dno akwarium w ten sposób, że ryby go nie widziały, odnajdywały go przeważnie dopiero po dotknięciu go jednym z nitkowatych promieni płetw piersiowych.
W sumie więc obserwacje wstępne nad rybami z rodziny Osphromenidae, głównie nad Trichogaster trichopterus var. sumatranus, wskazują, że zwierzęta używają swych nitkowatych wyrostków płetwowych do wyszukiwania pokarmu i to zarówno unoszącego się w wodzie, jak i spoczywającego na dnie.
Doświadczenia nad wrażliwością dotykową
Rola nitkowatych wyrostków płetwowych u badanych ryb mogła polegać na dodatkowym rozróżnianiu przedmiotów lub na chemicznym rozpoznawaniu substancji, lub na obu funkcjach równocześnie. W dalszej serii doświadczeń zajęto się najpierw zanalizowaniem wrażliwości dotykowej. О wrażliwości promieni płetwowych na bodźce mechaniczne świadczyły obserwacje nad sposobem zachowania się ryb w prawie zupełnych ciemnościach. Wyrostki te wyraźnie pomagały zwierzętom przy omijaniu przeszkód i informowały o zbliżaniu się obcych ciał. Ryby pływały zupełnie pewnie i swobodnie, trzymając szeroko rozstawione i wysunięte ku przodowi wyrostki i poruszając nimi we wszystkich kierunkach, jakby obmacując otoczenie. Omijały one w ten sposób nie tylko liście roślin wodnych znajdujące się w akwarium, ale także płytki szklane lub pręty poustawiane na ich drodze. Podobne zachowanie się obserwowałem wielokrotnie także u ryb pozbawionych wzroku, które miały zwyczaj nocowania zawsze w jednym miejscu. Gdy po opuszczeniu przez ryby miejsca noclegowego, przestawiono ułożenie liści walisnerii, nie powodowało to nigdy zaburzeń w powrocie na to samo miejsce. Ślepa ryba odnajdywała z łatwością niespodziewane przeszkody i omijała je skrzętnie, dotykając ich uprzednio wysuniętymi do przodu nitkowatymi promieniami płetwowymi.
Obserwowane w niniejszej pracy gatunki ryb w czasie spoczynku nocnego zawsze wystawiały nitkowate wyrostki do przodu i wykonywały nimi bardzo powolne ruchy. Nagłe dotknięcie nitkowatego wyrostka spoczywającej ryby pałeczką szklaną wywoływało nieodmiennie reakcję przestrachu i szybką ucieczkę. Reakcja ta była przy tym wyraźniejsza, im bliżej ciała następowało podrażnienie mechaniczne. Ryby karmione w ciemnościach wykonują ruchy chwytania pokarmu jedynie wtedy, gdy pokarm dotknie ich nitkowatych wyrostków płetwowych.
Wyniki tych obserwacji i doświadczeń wskazują, że wyrostki nitkowate ryb z rodziny Osphromenidae są wrażliwe na bodźce mechaniczne, przy czym na zadrażnienia tego typu nastawione są prawdopodobnie wolne zakończenia nerwowe.
Doświadczenia nad wrażliwością chemiczną
W dalszej części doświadczeń starano się zbadać, czy nitkowate promienie płetwowe ryb z rodziny Osphromenidae, oprócz wrażliwości dotykowej, posiadają także właściwości receptorów zmysłu chemicznego. Ażeby wyeliminować przy reakcjach wykonywanych przez zwierzęta ewentualny udział zmysłu wzroku, przeprowadzono także szereg prób na rybach ślepych.
Ryby pozbawione wzroku, po przejściu szoku pooperacyjnego, przyzwyczaiły się do nowych warunków i pływały omijając przeszkody, tak samo jak ryby posiadające normalnie rozwinięte oczy. Wyszukiwały one łatwo pokarm wrzucony na dno, macając po dnie akwarium swymi nitkowatymi promieniami płetwowymi. Ponieważ jednak końce nitkowatych wyrostków wyszukiwały pokarm dość daleko przed otworem gębowym, zdarzało się często, że gdy ryba zwróciła się pyszczkiem do pokarmu, pokarm ten przesuwał się pod nią i nie został pobrany. Reakcja zwracania się do pokarmu była u ryb ślepych tak szybka, że przeważnie potrafiły one schwycić pokarm opadający powoli na dno. Nie potrafiły one jednak nigdy pochwycić dotkniętych nitkowatymi wyrostkami rozwielitek (Daphnia).
Dla zbadania wrażliwości płetwowych wyrostków nitkowatych na bodźce chemiczne przeprowadzono następujące serie doświadczeń.
1. Rybom ślepym i nieco wygłodzonym podano najpierw wazonkowce (Enchytreus) zawieszone na cienkim drucie. Aż do czasu zjedzenia dostatecznej ilości pokarmu ryby okazywały reakcje zestawione na tabelach 2 i 3. Z zestawień tych wynika, że ryby w znacznym procencie okazują reakcje dodatnie na podawane im wazonkowce, kierując się głównie zmysłem chemicznym umiejscowionym na nitkowatych wyrostkach płetwowych. Przy wyszukiwaniu pokarmu musi jednak odgrywać także rolę dodatkową i wzrok. Wynika to z porównania tabeli 1 i tabel 2 i 3.
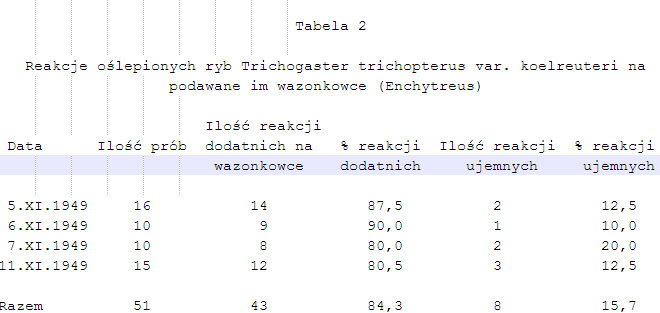
Ryby z normalnymi oczami reagują szybko i pewnie w 100%. Ryby ślepe natomiast wykazują już tylko 84,3% lub 87,5% reakcji dodatnich. Jak wspomniano wyżej, doświadczenia z rybami o normalnym wzroku były przeprowadzane w ten sposób, że ryba w chwili podawania pokarmu nie mogła go widzieć. Pierwszy sygnał o jego obecności pochodził od nitkowatego wyrostka płetwowego, który się z nim zetknął. Sygnał ten mógł być dotykowy lub chemiczny. Dopiero później ryba zobaczyła pokarm i gdy wzrok upewnił ją o jego obecności, następowało 100% reakcji dodatnich. Ryba ślepa nie mogła posługiwać się wzrokiem. Pierwsze sygnały o obecności pokarmu pochodziły również od nitkowatych wyrostków, które odbierały odnośne podniety dotykowe i chemiczne. Podniety te same były jednak tu za słabe, aby ryba nabrała od razu w 100% pewności, czy podniety otrzymywane za pomocą nitkowatych wyrostków informują ją o pokarmie, czy o przedmiocie dla niej obojętnym.
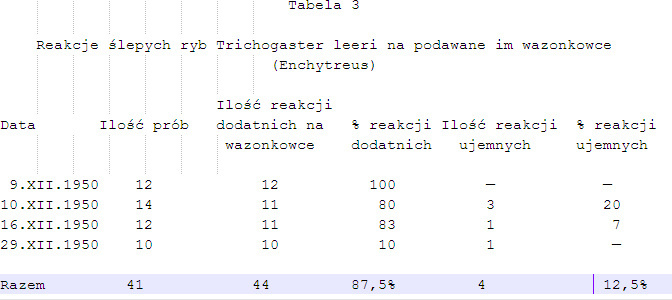
2. Dla dokładniejszego zbadania, czy reakcje ryb oparte są istotnie na wrażliwości chemicznej wykonano dalsze doświadczenia, w których rybom ślepym podawano do wyboru wazonkowce (Enchytreus) i kawałki nitek bawełnianych odpowiedniej wielkości przepojone sokiem uzyskanym z rozgniecionych wazonkowców. Wyniki odnośnych prób zestawione zostały w tabeli 4. Jak wynika z procentów dodatnich reakcji na żywe wazonkowce, i nitki przepojone ich sokiem, odnośne wartości stoją bardzo blisko siebie. Podkreślić przy tym należy, że ryby głodne reagowały początkowo prawie jednakowo na oba rodzaje podawanych im przynęt. W miarę jednak powtarzania doświadczeń procent dodatnich reakcji na żywe wazonkowce zwiększył się i stale przewyższał procent reakcji dodatnich na nitkę przepojoną sokiem z wazonkowców. Wynika stąd wniosek, że choć podniety chemiczne są jednakowe w obydwu przypadkach, to jednak żywy, poruszający się energicznie wazonkowiec (Enchytreus) działa widocznie na ryby silniej pociągająco niż nitka. Wydaje się także, że podniety chemiczne działają przy tym słabiej niż podniety dotykowe. Uzyskany w tej serii doświadczeń (tabela 4) procent reakcji ujemnych, w przybliżeniu taki sam jak przy podawaniu rybom samych wazonkowców w poprzedniej serii doświadczeń (tabela 2 i 3), również zdaje się wskazywać na pewną rolę jaką przy poszukiwaniu pokarmu w warunkach normalnych odgrywa wzrok.
3. Podobne doświadczenia powtórzono jeszcze raz z tym, że zamiast wazonkowców (Enchytreus) użyto do karmienia ryb mięsa, przy czym karmiono je przez dłuższy czas. Przy użyciu mięsa końskiego wyniki badań pokrywały się z wynikami uzyskanymi przy użyciu wazonkowców. Przy użyciu mięsa innego, np. wieprzowiny, baraniny lub mięsa rybiego, zwierzęta doświadczalne brały pokarm tylko na początku prób, a następnie bardzo szybko przestawały reagować nie tylko na nitki przepojone sokiem
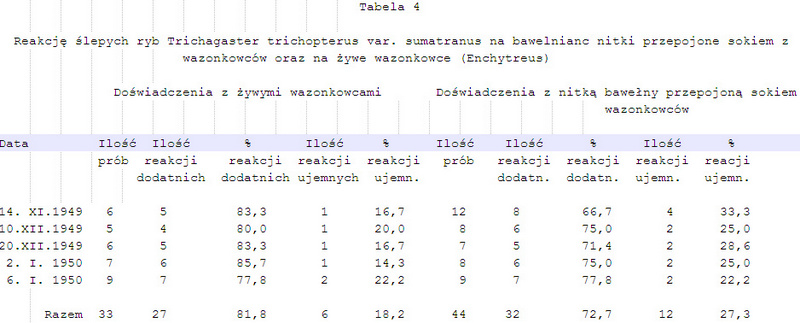
z tych mięs, ale także na samo mięso odpowiedniej kategorii, a niekiedy nawet okazywały wyraźne reakcje ucieczki. Fakt ten można by wytłumaczyć albo tym, że zwierzęta nie znały jeszcze substancji chemicznych pochodzących z tych mięs, albo że nie działały one podniecająco na ryby w kierunku reakcji chwytania pokarmu. Substancje te mogły nawet w pewnych warunkach działać odpychająco, wywołując reakcje ucieczkowe. Ostatni fakt byłby zgodny z wynikami badań v. Frischa (1938), który wyraźne reakcje ucieczkowe na krew ryb stwierdził u strzebli (Phoxinus laevis).
Ogólnie biorąc, z doświadczeń powyższych wynika, że ryby z badanych gatunków odbierają przy pomocy nitkowatych promieni płetwowych nie tylko podniety mechaniczno-dotykowe, lecz także i chemiczne. Te ostatnie działają szczególnie wówczas, gdy widzialność przedmiotów w zanieczyszczonej wodzie jest niewielka. Ażeby sprawdzić wniosek ten przeprowadzono jeszcze następujące doświadczenie.
4. Rybom pozbawionym wzroku, karmionym wazonkowcami (Enchytreus), podawano do wyboru wazonkowce i nitki bawełniane nie nasycone sokiem z tych pierścienic. Początkowo ryby chwytały jedne i drugie bez wyboru, szybko jednak nauczyły się odróżniać wazonkowce od nitek bawełnianych. W ostatecznym wyniku ilość reakcji na te ostatnie bardzo zmalała. Wyniki te zostały przedstawione na tabeli 5. Wymaga ona dodatkowych objaśnień. Jak to podkreślono wyżej, reakcje ujemne odnoszą się zwykle do początku prób. Znaczny procent reakcji ujemnych, bo aż 28,6%, zanotowany w dniu 17.IV.1950 prawdopodobnie był wynikiem przegłodzenia ryb i przez to dążenia ich do jak najrychlejszego zaspokojenia głodu bez zwracania specjalnej uwagi na jakość podawanych im wabików. W doświadczeniach tych stwierdzono również, że wystarczył

jeden dzień, aby ryby popełniały na początku nowej serii prób pomyłki. Mimo wielokrotnych usiłowań, nie udało się doprowadzić do tego, aby ryby od razu brały pokarm bezbłędnie.
Podobne wyniki uzyskano przy stosowaniu zamiast wazonkowców, pokarmów innych, np. mięsa końskiego. Doświadczenia tej serii wskazują również, że ryby doświadczalne kierują się przy wyszukiwaniu pokarmu równocześnie zmysłem dotyku i zmysłem chemicznym. Podniety chemiczne muszą przy tym działać znacznie słabiej, albowiem zwierzęta zapominają je szybciej.
5. W dalszych seriach badań próbowano działać na nitkowate wyrostki płetwowe ryb niektórymi charakterystycznymi substancjami chemicznymi działającymi na zmysł smaku, głównie kwasem solnym, oraz roztworami chininy i cukru. Doświadczenia przeprowadzono w ten sposób, że na nitkowaty wyrostek puszczano z pipety delikatny strumień odnośnego związku chemicznego. Reakcje ryb w tym przypadku nie były charakterystyczne. Po pewnej chwili od wypuszczenia danego roztworu ryby wykazywały niepokój i uciekały, ale reakcja ta mogła równie dobrze pochodzić stąd, że po pewnym czasie płyn wpuszczony do wody mógł działać nie tylko na receptory zmysłu chemicznego umieszczone na nitkowatych wyrostkach ale także na receptory mieszczące się w pobliżu otworu gębowego i w jamie gębowej. Кwas solny mógł poza tym działać parząco.
6. Przypuszczenia te zdają się potwierdzać dalsze doświadczenia kontrolne przeprowadzone z jednej strony na rybach o normalnie rozwiniętym wzroku, z drugiej na rybach ślepych. Zwierzętom podawano pokarm sztuczny, tzw. „Wawil”. Część jego moczona była w roztworze chininy i dla odróżnienia wzrokowego dla obserwatora od pokarmu czystego zabarwiona błękitem metylenowym, druga zaś część niezaprawiona chininą pozostawała niezabarwiona. Wyniki wyborów obu rodzajów pokarmu sztucznego przedstawiono na tabeli 6. Wynika z nich, że w niektóre dni ryby nie brały zupełnie pokarmu sztucznego zaprawionego roztworem chininy, w inne zaś procent reakcji chwytowych na pokarm taki był znikomy, najwyżej do 7,7%. Zdecydowana większość wyborów przypadała zawsze na pokarm sztuczny niczym nie zaprawiony. Podkreślić należy, że przy karmieniu „Wawilem” ryby zawsze próbowały go przy pomocy nitkowatych wyrostków płetwowych. Przy próbach z rybami о normalnie rozwiniętych oczach reakcje ujemne następowały dopiero wówczas, gdy zwierzęta wyjadły zupełnie pokarm sztuczny niczym nie zaprawiony. Natomiast po małej ilości chwytów za „Wawil” zaprawiony chininą i barwikiem ryby przestawały reagować na niego pozytywnie, mimo że dotykały go wyrostkami nitkowatymi. Również pokarm sztuczny normalny pływający zaprawiony chininą i barwikiem długi czas pozostawał nietknięty. W analogicznych doświadczeniach z rybami pozbawionymi wzroku,
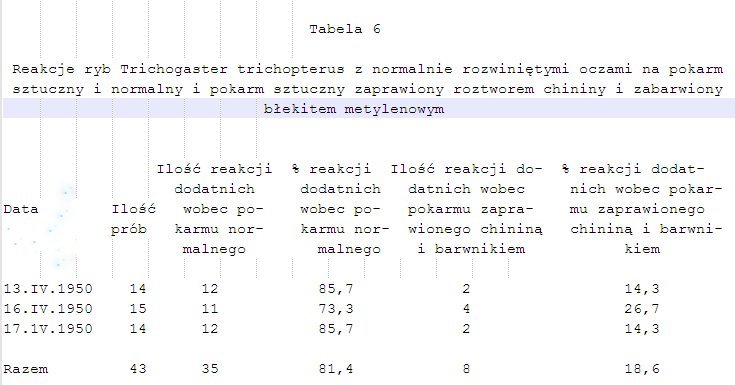

których wyniki przedstawiono na tabeli 7, procent reakcji chwytowych na pokarm normalny jest bardzo duży, mniejszy jednak niż u ryb z normalnie rozwiniętym wzrokiem. Procent reakcji na pokarm zaprawiony chininą niewiele odbiega od procentów reakcji ujemnych uzyskanych przy innych, omówionych już poprzednio, doświadczeniach. Wszystkie te fakty potwierdzają znów przypuszczenie, że przy wyszukiwaniu przez badane ryby pokarmu, obok bodźców dotykowych i chemicznych, odbieranych przy pomocy receptorów umieszczonych na nitkowatych promieniach płetwowych, pewną rolę odgrywa także wzrok. Ryba pozbawiona możności sprawdzenia wzrokiem podniet dochodzących z nitkowatych wyrostków, jest mniej pewna siebie przy wyszukiwaniu pokarmu i łatwiej popełnia błędy.
Doświadczenia dodatkowe nad rolą wzroku
przy wyszukiwaniu pokarmu przez ryby
Na rybach o normalnie rozwiniętych oczach i rybach ślepych wykonano jeszcze następujące serie doświadczeń. Do akwarium z rybami normalnie widzącymi wkładano dość głęboko pod powierzchnię wody pionowo probówkę, w której umieszczono drobno posiekane kawałeczki mięsa. Ryba zobaczywszy pokarm umieszczony na dnie probówki, przez szklane jej ściany, nie atakowała go bezpośrednio pyszczkiem, lecz starała się zbadać go, obmacując probówkę nitkowatymi wyrostkami na tej wysokości, na której znajdowało się mięso. Po krótkim czasie jej zainteresowanie wzrokowe pokarmem słabło, natomiast ryba, nie przestając dotykać nitkowatymi wyrostkami probówki, posuwała się coraz wyżej. W miarę jej wznoszenia się wzdłuż probówki, wzrastała też szybkość jej przemieszczania się w górę, wreszcie u brzegu otwartego probówki następował bardzo szybki atak ryby na brzeg probówki i chwyt pyszczkiem.
Wielokrotne próby wykazały zupełnie taki sam sposób zachowania się ryb. U ryb ślepych, którym probówkę z ukrytym w niej na dnie pokarmem, trzeba było podstawić całkiem blisko, reakcje przebiegały nieco inaczej. Obmacywały one probówkę, wykonując reakcji chwytowych przez pewien czas i przesuwając się niepewnie to w górę, to w dół. Po pewnym czasie ryba podpływała zdecydowanie wzdłuż probówki do jej otworu, gdzie dopiero następował chwyt pyszczkiem za pokarmem znajdującym się w probówce. Przebieg tych doświadczeń można wytłumaczyć w następujący sposób. U ryb z normalnie rozwiniętym wzrokiem, ten ostatni, jak widać było z poprzednich doświadczeń, odgrywa pewną rolę przy odszukiwaniu pokarmu umieszczonego w probówce. Stąd pochodził natychmiastowy zwrot ryby w kierunku pokarmu umieszczonego w probówce. Gatunki ryb, na których prowadzono badania, tylko wyjątkowo chwytają pokarm bez dotknięcia go swymi nitkowatymi wyrostkami płetwowymi. Dlatego też dotykały one probówkę na wysokości znajdującego się w niej pokarmu. Ponieważ wyrostki nitkowate napotykały na szkło i ryba początkowo nie odczuwała żadnych podniet chemicznych, przeto nie atakowała pokarmu. Po pewnym czasie jednak substancje chemiczne wydzielające się z mięsa, dyfundowały do wody otaczającej probówkę. Stężenie ich było naturalnie najsilniejsze przy otworze probówki i malało w miarę wzrastania odległości od tego otworu. Substancje te, działając prawdopodobnie na receptory zmysłu chemicznego mieszczące się na nitkowatych wyrostkach, powodowały odpowiednią reakcje ryby, polegającą na przesuwaniu się jej wzdłuż probówki stosownie do gradientu stężenia. Gdy zwierzę dostało się w ten sposób w pobliże otworu probówki, gdzie koncentracja substancji chemicznych pochodzących z mięsa znajdującego się na dnie probówki była największa, i już więcej nie wzrastała, zwierzę mogło przypuszczać, że znajduje się w bezpośredniej bliskości pokarmu, co powodowało chwyt za brzeg probówki.
Przebieg doświadczeń wskazuje, że zmysł wzroku odgrywał rolę na samym początku. Później zwierzę poczuwszy zmysłem chemicznym substancje dyfundujące z otworu probówki, przestało zwracać uwagę na widok pokarmu a skierowało się ku największym stężeniom substancji pochodzących z pokarmu. Potwierdzają to również doświadczenia z rybami pozbawionymi wzroku. Jak wspomniano wyżej, początkowo obmacywały one probówkę w różnych miejscach, a nie od razu na wysokości, gdzie znajdował się pokarm, którego nie mogły widzieć. Z chwilą jednak, gdy dotarły do nich pierwsze ślady substancji mięsnej dyfundujące z otworu probówki, zaczynały płynąc zdecydowanie do otworu, gdzie następował podobny chwyt za brzeg probówki.
Zebranie wyników
Przedstawione doświadczenia nad reakcjami ryb z rodziny Osphromenidae nad wyszukaniem pożywienia pozwalają stwierdzić, że w pierwszym rzędzie kierują się one wzrokiem. Dostrzegłszy jakieś ciało, które mogłoby stanowić pożywienie, zbliżają się do niego i przy pomocy długich nitkowatych, wolnych promieni płetw piersiowych zaczynają je badać dokładniej. Jeżeli ciało to odpowiada im, chwytają je pyszczkiem i połykają. Ryby posiadające normalny wzrok reagują na rozmaite ciała podawane im szybciej i pewniej niż ryby ślepe. Po wyłączeniu wzroku ryby w poszukiwaniu pokarmu kierować się mogą wyłącznie samą wrażliwością nitkowatych wyrostków płetwowych. Pominięte zostały tutaj naturalnie dla jaśniejszego obrazu inne narządy zmysłowe umieszczone na innych częściach ciała. Wyrostki te spełniają podwójną funkcję narządów dotykowych i narządów chemicznych. O pierwszej funkcji świadczy fakt obmacywania w czasie płynięcia naprzód przestrzeni znajdującej się przed nimi, aż chwilą zauważenia jakiegoś przedmiotu dotykania go z różnych stron. Funkcje receptorów chemicznych wykazały doświadczenia z rozróżnianiem przez ryby substancji jadalnych od niejadalnych lub substancji jadalnych normalnych i zaprawianych chininą.
Doświadczenia prowadzone nad kilkoma gatunkami ryb z rodzajów Colisa i Trichopteus z rodziny Osphromenidae, wykazały, że reakcje przy pomocy wyrostków nitkowatych na bodźce dotykowe, jako i chemiczne, są u różnych gatunków podobne.
Wyniki uzyskane nad rybami z rodzaju Colisa i Trichogaster z rodziny Osphromenidae dotyczące funkcji dotykowej i chemicznej nitkowatych wolnych promieni płetw piersiowych wykazują dużą zgodność z wynikami uzyskanymi odnośnie funkcji zmysłowej podobnych wyrostków u innych ryb, a przede wszystkim u przedstawicieli rodziny Triglidae. Świadczy to o wykształceniu analogicznych narządów zmysłowych na homologicznych utworach ciała, w tym przypadku wolnych promieniach płetwowych, w zależności od podobnego trybu życia i poszukiwania pokarmu.
PIŚMIENNICTWO
1. Allen H. (1855). On the pectoral filaments in the sea rober (Prionotus palmipes). Proc. Acad. Natur Sci. Philad. 377.
2. Dykgraaf S. (1934). Untersuchungen über die Funktion der Seitenorgane an Fischen. Ztschr. f. vergl. Physiol. 20. 162—214.
3. d’Evant T. (1903). L’epitelio sensitivo del raggi digitali delle Trigle. Giorn. Assoc. Napol. Med. e Natural. 13. 3—29.
4. v. Frisch K. (1926). Vergleichende Physiologic des Geruchs- und Geschmacksinnes. Hdb. d. norm. u. patol. Physiol. 11.
5. v. Frisch K. (1938). Zur Psychologie des Fisch-Schwarmes. Naturwiss. 26.
6. Herrick C. J (1903). On the morphological and physiological classification of the cutaneous sense organs of fishes. Amer. Nat. 37. 313—318.
7. Herrick C. J. (1904). The organ and sense of taste in fishes. Bull. US Fish Comm. Washington 22, 239—272.
8. Herrick C. J. (1907). The tactile centres in the spinal cord and brain of the sea robin Prionotus carolinensis L. J. comp. Neur. 17, 307—327.
9. Jourdan E. (1890). Structure histologiue des barbillons et des rayons libres du Peristedion cataphractum. Arch, de Zool. II. 8, 603—616.
10. Krinner M. (1935). Über die (Geschmacksempfindlichkeit der Elritzen. Ztschr. f. vergl. Physiol. 21. 317—342.
11. Morril A. D. (1849). The pectoral appendages of Prionotus and their innervation J. Morph. a. Physiol. 11, 177—192.
12. Nagel W. A. (1894). Vergleichend Vergleichend physiologische und anatomische Untersuchungeu über den Geruchs- und Geschmackssinn und ihre Organe. Bibliotheca. zool. 7. zesz. 18, 207.
13. Olmsted J. M. D. (1920). The results of cutting the seventh cranial nerve in Amiurus nebulosus (Lesueur). J. Exp. Zool. 31, 369—401.
14. Parker G. H. (1908). Sense of taste in fisches. Science US 27, 453.
15. Parker G. H. (1910). Olfactory reactions in fishes. J. Exp. Zool. 8, 535—542.
16. Parker G. H. (1922). Smell, taste, and allied senses in the vertebrates. Philadelphia, Lippincott, 1—192.
17. Piéron H. (1908). Contribution à l’étude des phénomena inférieurs. Bull. Inst, gen. psychol. 8, 321—327.
18. Scharrer E. (1935). Die Empfiudlichkeit der freien Flossensstrahlen des Knurrhans (Trigla) fur chemische Reize. Ztschr. f. vergl. Physiol. 22, 145—154.
19. Sheldon R. E. (1909). The reactions of the dogfish to chemical stimuli J. comp. Neurol. 91, 273—311.
20. Sheldon R. E. (1911). The sense of smell in selachians. J. Exp. Zool. 10, 51—62.
21. Steiner J. (1885). Die Funktionen des Centralnervensystems und ihre Phylogenese. Braunschweig, Vieweg. 1—127.
22. Strieck R. (1924). Untersuchungen über den Geruchs- und Geschmackssinn der Erlitzen. Ztschr. f. verg. Physiol. 2, 122—154.
23. Tiedemann F. (1816). Von dem Hirn und fingerförmigen Fortsätzen der Triglen. Dfsch. Arch. fd Physiol. 2. 103—110.
24. Uexküll J. v. (1895). Vergleichend-sinnesphysiologische Untersuchngen. Z. Biol. 32, 549—566.
25. Ussow M. (1882). De la structure des lobes accessories de la moelle épinière de quelques poissons osseux. Arch. d. Biol. 3, 605 658.
26. Warden C. J., Jenkins T. N., Warner L. H. (1936). Comparative psychology. A comprchensive treatise. III, Vertebrates. New York, The Ronald Press Pomp.
SUMMARY
Experiments on the sensory functions of the pectoral filaments of the pectoral fins of fishes of the family Osphromenidae were made on the following species: Colisa fasciata Bloch & Schneider, C. labiosa Day, Trichogaster leeri Blecker., Tr. trichopterus Pall. var. koelreuteri Cuv. & Val., and Tr. trichopterus Pall. var. sumatranus Ladiges. From the observations and experiments shown in tables 1 — 7 it results that these fishes, irrespectively of the species, only take food after touching it with the said pectoral filaments playing a double part of feeling and chemical organs. The feeling sensitiveness of the free pectoral filaments allows the fishes to avoid obstacles in the dark, or blind fishes to orientate at all. Thanks to the sensitiveness of these filaments to chemical incentives the fishes may discern bodies suitable as food from other ones. Sight also plays a part in seeking food. Blind fishes recognise food thanks to the chemical sense. When they were given live specimens of Enchytreus and cotton threads imbibed with juice from Enchytreus or untreated ones, the fishes initially took both; however, they gradually learnt to distinguish both baits and mostly took the live Enchytrei avoiding the cotton threads. Similar results were reached with meat of different kinds served as food. Sometimes the fishes fled from meat. The fishes also recognised with their pectoral filaments artificial food from a similar one flavoured their pectoral filaments artificial food from a similar one flavoured with quinine and avoided the latter. Experiments with food placed in test- tubes showed the additional role of sight in seeking food. Fishes with normal eyes felt the test-tube outwardly in the place where the food was placed, while the blind fishes felt the test-tubes in various places and showed seizing reactions only when the substances from the food began to diffuse into the surrounding water. The feeling and chemical sensory functions of the free pectoral filaments of fins in fishes of the family Osphromenidae are similar to the functions of similar organs in fishes of the family Triglidae.